




Addition of functionality to a bacterial genome: added information
Alec MacAndrew
Introduction
The genome of Streptomyces coelicolor has recently been fully sequenced at the Sanger centre in the UK, Nature 417, 141 -147, Bentley et al (1)S coelicolor is a soil bacterium that has many different metabolic processes and biotransformations that allow it to live under a wide range of conditions in the soil and use a wide range of metabolic pathways to, for example, degrade insoluble remains of other organisms. S coelicolor is unusual, in that it is a multicellular bacterium which forms into different sorts of 'tissues'. The complexity of its metabolic pathways give rise to two thirds of modern antibiotics, as well as anti-tumour and immuno-suppressant agents. All this complexity arises from the longest known eubacterial genome (8.7 million base pairs and 7,825 genes; about one quarter the number of human genes).
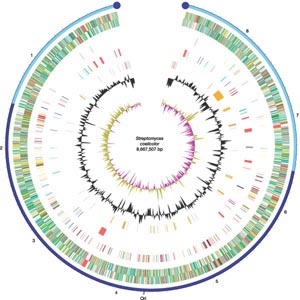
The S coelicolor genome
The S coelicolor
chromosome is linear. It is divided into a core
region which extends from 1.5Mb to 6.4Mb (a length of 4.9Mb or million letters)(1.5Mb to 6.4Mb
means megabases or millions
of genetic letters - this is a position on the chromosome - we start
counting at one end which is position zero; the other end is position 8.7
million) ; and into two other
regions, one before 1.5Mb on the leading arm and one after 6.4Mb on the
trailing arm. All of the genes essential for the viability of the organism
lie within the core region. That means that the essential-for-life genes are
clustered into the central region. Genes associated with secondary
(non-essential function, or nice-to-have) lie in the outer arms.
S
coelicolor is a actinomycete and so are Mycobacterium tuberculosis and
Corynebacterium diptheriae (these are the bacteria that cause diphtheria and
tuberculosis respectively). The genomes of these bacteria are respectively about 4Mb and 2.5Mb long.
The core of S coelicolor (that is the central region between 1.5Mb and
6.4Mb) and the entire genomes of M tuberculosis and C diptheriae (that means
all their chromosomes) show considerable similarity at the level of
individual gene sequences - they have very many genes which show a great
deal of similarity. Even more telling, all three genomes (the core of S
coelicolor and the entire genomes of M tuberculosis and C diptheriae) show
considerable synteny - that means that not only are the same genes present
but they are in the same sequence on the genome - they are arranged in
the same order. It is therefore likely that the core of S coelicolor and
the entire genomes of the other two bacteria have a common
ancestor.
However, the genes outside the core region of S coelicolor
(in those arms before 1.5Mb and after 6.4Mb) show no synteny (genes in the
same order) with the other bacteria. So for the following reasons, in S
coelicolor, the leading arm up to 1.5Mb and the trailing arm after 6.4Mb
are later additions to the genome:
1) These regions show no synteny
(genes in the same order) with related genomes of M tuberculosis and C
diphtheriae
2) These regions do not contain genes essential to the viability of the organism
3) These regions can undergo extensive deletions (removal of genetic material) or amplifications (addition of genetic material) without affecting the viability of the organism
4) Many of the genes in
these regions are associated with regulation, transport and degradation of
extracellular nutrients and are not associated with basic
viability.
So where did these additional genes in the arms of the
chromosome of S coelicolor come from? 14 sequences in the arms correlate
with known mobile genetic elements and are therefore likely to be Insertion
Sequences. (There are
certain genetic sequences which code within themselves the ability to move
from the chromosome of one organism to another - even where the
organisms are different species). Six others are the same as plasmid
genes which are found in the related bacterium S ambofaciens pSAM2
plasmid. (Plasmids are cellular structures which contain separate islands of DNA - genetic material -
found in bacteria separate from the chromosome. There is a lot of
evidence that in evolution genes can migrate from bacterial plasmids to
bacterial chromosomes) There is also evidence for transposition within the
chromosome and gene duplication.
So in this bacterium, an original core
chromosome has had an additional 3.8Mb of DNA added in evolutionary
development which has resulted in a morphology (morphology means the
physical form the bacterium takes - non-dividing many branched mycelium
versus dividing cells) and a metabolism (how it gets its food and creates
its energy for living) that is more complex and very different from its
relatives.
There is a huge amount of functionality that has been added
to the arms of the chromosome through the evolutionary process: new genes
coding for new functions.
Conclusion
So we see that functionality and DNA has been added to genome of S coelicolor since its divergence from M tuberculosis and C diphtheriae; evidence which stand in contradiction to the claim that information cannot be added to the genome through evolution.
1. Bentley et al, Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2), Nature 417, 141 -147
